How EPAX® Omega 3-9-11 may support a healthy inflammatory response


EPAX® Omega 3-9-11 combines marine Omega-3s EPA and DHA with long-chain monounsaturated fatty acids, including cetoleic acid and gondoic acid. Emerging research suggests this unique composition may support a healthy inflammatory response, with particular relevance for adipose tissue, metabolic health, and skin health.
Composition
EPAX Omega 3-9-11 is a concentrated fish oil with minimum requirement of long chain PUFA and long chain MUFAs as shown in Table 1.

Independent investigator driven studies most often pose the question of the specific role of LCMUFAs and therefore require an oil with Omega-9 and Omega-11 and low levels of EPA and DHA, and a placebo with an equal amount of Omega-3 fatty acids. These ask the basic question: Do the LCMUFAs provide a desired effect.
Hence, the majority of studies investigating mechanism of action favour use of a cetoleic acid concentration with low EPA and DHA content.
Biodistribution
LCMUFAs and EPA/DHA may have overlapping target molecules, for example PPAR, although the physiological outcome may be different. One hypothesis for this difference lies in the biodistribution of EPA/DHA contra LCMUFAs.
Pre-clinical studies show that LCMUFAs accumulate to a high degree in adipose tissue and less in other tissues. EPA/DHA has the highest accumulation in the brain and retina and lowest in adipose tissue amongst tissue tested.
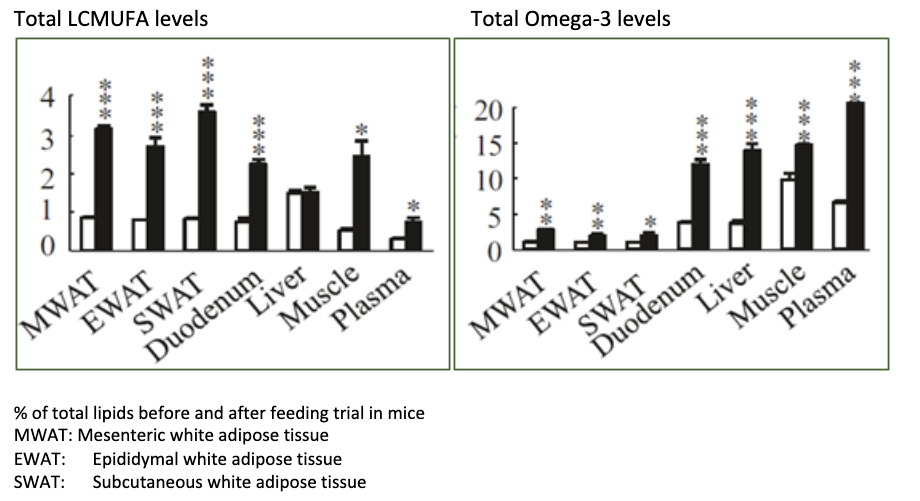
Yang et al hypothesise that this differential distribution of the fatty acids may lead to different effects on the body [1]. This can be of particular significance in metabolic health where low-grade chronic inflammation of adipose tissue is seen in overweight/obesity which impacts glucose metabolism, insulin sensitivity and circulating lipids.
In diabetic mice, LCMUFA supplementation reduced skin fat content by 16%, without changes in food intake or body weight [2]. This strongly supports a unique and targeted activity of LCMUFAs on adipose tissue.
Preclinical data on inflammation
Pre-clinical studies by Yang et al., at the NIH demonstrate that mice fed an LCMUFA diet have increased expression of the anti-inflammatory signalling molecule PPAR(2). In the same study, diet induced obese mice showed significant reductions in TNF-alpha and matrix metalloproteinases with an LCMUFA diet which was also shown in additional pre-clinical studies (3, 4).

Adiponectin is a signalling molecule released by adipocytes with autocrine and endocrine activity. This molecule has a number of down-stream effects including anti-inflammatory activity and systemic metabolism. Adiponectin levels were improved in several pre-clinical studies following an LCMUFA diet (5, 6).
Studies with EPAX Cetoleic concentrate
Pre-clinical studies were performed in rats using an Epax cetoleic concentrate or placebo with balanced EPA/DHA. These studies showed the cetoleic concentrate reduced cholesterol levels in a diabetic rat model (7) and an obesity rat model (8).
Supporting these was a third study assessing inflammation in an obesity rat model (9).
Feeding with EPAX Cetoleic acid concentrate led to significant reductions in inflammatory biomarkers in white adipose tissue, after 35 days of feeding.
Reductions were dose dependent with a low-dose natural oil giving reduced benefit compared to the concentrate. The concentrate had approximately double the amount of LCMUFA compared to the natural oil.
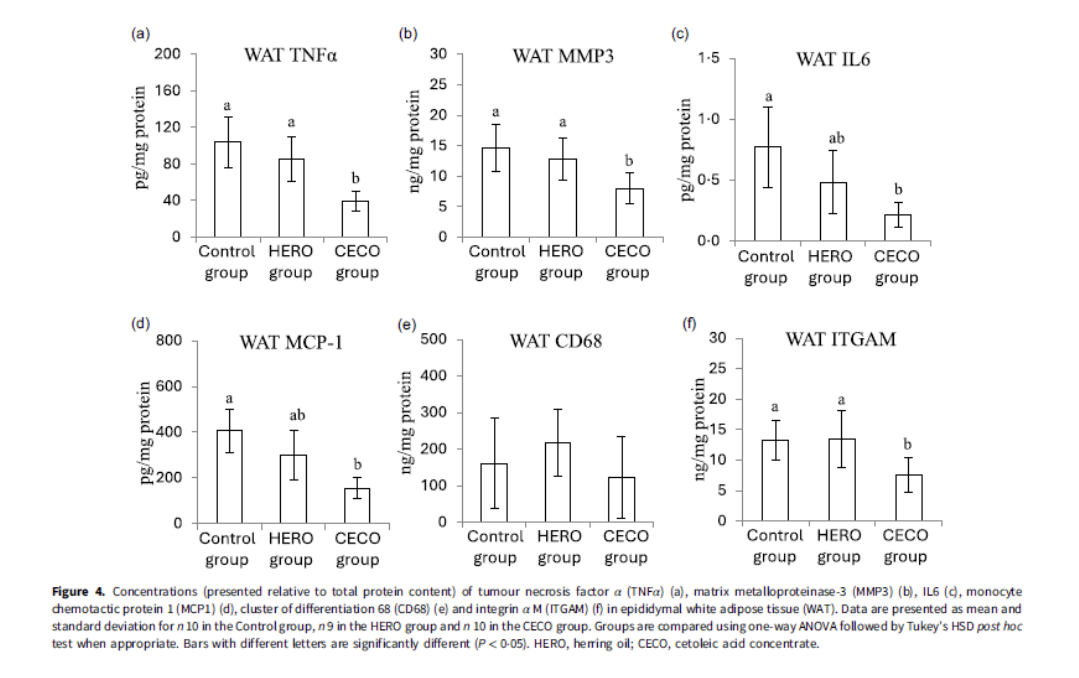
Figure 3 shows significant reductions in TNF-α, a master regulator of inflammation, Il-6 an inflammatory cytokine, MMP3 a matrix metalloproteinase important for tissue re-modelling but also with a role in cancer metastasis and collagen breakdown, MCP-1 /CD68/ITGAM which are all immune function receptors.
Interestingly, the in vivo data above is supported by cell-culture data from psoriasis research performed by the Norwegian research institute NOFIMA (10). Here, the research was targeted towards psoriasis-relevant inflammatory biomarkers.
In keratinocytes stimulated with TNF-α and IL-17 the production of the pro-inflammatory IL-6 was unaffected by co-incubation with DHA or EPA but significantly reduced with synthetic cetoleic acid (labelled as Ketolinsyre).
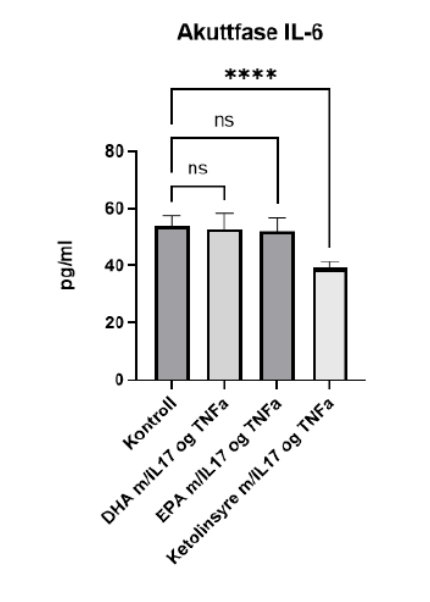
The study goes on to show that cetoleic acid reduces inflammation (IL-8) in adipocytes and improves adipocyte mitochondrial function. In the model system used in this study, (ie IL17 stimulated inflammation) EPA and DHA had effects on resolution of inflammation and cetoleic acid on the initial inflammatory response. The authors conclude by proposing a potential synergistic effect of combined Omega-3 and long-chain monounsaturates in reducing the overall inflammatory process with this particular model system, as shown in Figure 5.
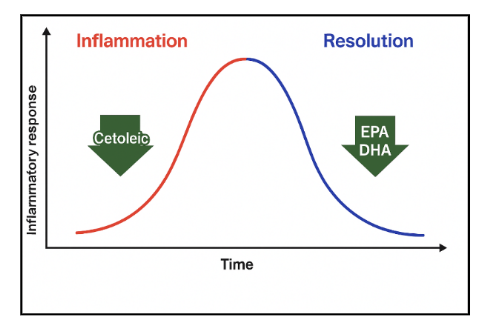
Omega-3 has broad anti-inflammatory effects
The purpose of this white paper is to summarise research with long-chain monounsaturated fatty acids, particularly C22 cetoleic acid and C20 gondoic acid.
Briefly, the omega-3 fatty acids EPA and DHA, have multiple anti-inflammatory actions. These include the disruption of the omega-6 eicosanoid pathway, the promotion of resolution through production of Specialised Pro-Resolving mediators (SPMs) [11], as precursors to endocannabinoids, through receptor mediated down-regulation of master controllers (PPAR, TNF-α and NF-kB), and macrophage switching from M1 to M2 phenotype.
Conclusion
Current research supports the action of cetoleic acid as an anti-inflammatory agent with particular effect in adipocytes which show high accumulation of this fatty acid when given as oral supplementation. Differential biodistribution of LC-PUFAs (EPA/DHA) and LC-MUFAs (cetoleic and gondoic acid) and biological effects supports a potential synergistic anti-inflammatory effect of long chain PUFAs and long chain MUFAs.
References
1. Yang, Z.H., et al., Long-term dietary supplementation with saury oil attenuates metabolic abnormalities in mice fed a high-fat diet: combined beneficial effect of omega-3 fatty acids and long-chain monounsaturated fatty acids. Lipids Health Dis, 2015. 14: p. 155.
2. Yang, Z.H., et al., Dietary supplementation with long-chain monounsaturated fatty acids attenuates obesity-related metabolic dysfunction and increases expression of PPAR gamma in adipose tissue in type 2 diabetic KK-Ay mice. Nutr Metab (Lond), 2013. 10(1): p. 16.
3. Tsutsumi, R., et al., Long-chain monounsaturated fatty acids improve endothelial function with altering microbial flora. Transl Res, 2021. 237: p. 16-30.
4. Yang, Z.H., B. Emma-Okon, and A.T. Remaley, Dietary marine-derived long-chain monounsaturated fatty acids and cardiovascular disease risk: a mini review. Lipids Health Dis, 2016. 15(1): p. 201.
5. Yang, Z.H., et al., Pollock oil supplementation modulates hyperlipidemia and ameliorates hepatic steatosis in mice fed a high-fat diet. Lipids Health Dis, 2011. 10: p. 189.
6. Yang, Z.H., et al., Beneficial effects of dietary fish-oil-derived monounsaturated fatty acids on metabolic syndrome risk factors and insulin resistance in mice. J Agric Food Chem, 2011. 59(13): p. 7482-9.
7. Rimmen, E.V., et al., Absorption, accumulation and metabolism of cetoleic acid from dietary herring oil in tissues of diabetic male Zucker Diabetic Sprague Dawley rats. Br J Nutr, 2025: p. 1-24.
8. Oldernes, H., et al., Intake of herring oils containing cetoleic acid resulted in a lower serum cholesterol concentration in male obese Zucker fa/fa rats. Br J Nutr, 2026: p. 1-10.
9. Hansen, A., et al., Intake of a cetoleic acid concentrate lowered concentrations of markers of inflammation and macrophage infiltration but did probably not increase EPA biosynthesis in male obese Zucker fa/fa rats. Br J Nutr, 2025. 134(11): p. 881-891.
10. Mona E. Pedersen, M.B., Bente Ruyter, Jesmond Dalli, Achilleas Karvounopoulos, Torsetein Tengs, Nina Solberg, Vibeke Høst, Vibeke Voldvik, Astrid Nilsson og Tone-Kari K. Østbye., Effekt av ketolinsyre på psoriasis. 2026.
11. Spite, M., J. Clària, and C.N. Serhan, Resolvins, specialized proresolving lipid mediators, and their potential roles in metabolic diseases. Cell Metab, 2014. 19(1): p. 21-36.



